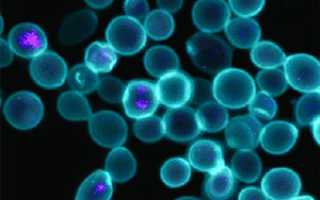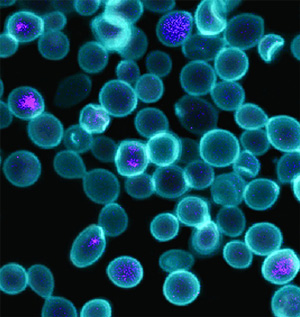
Рис. 1. Снимок химерных клеток кишечной палочки и дрожжей, сделанный с помощью конфокального флуоресцентного микроскопа. Голубым окрашены клетки дрожжей, фиолетовым — РНК кишечной палочки. Изображение из обсуждаемой статьи в PNAS
Общепринятая на данный момент теория симбиогенеза предполагает, что митохондрии в эукариотических клетках произошли от симбиотических бактерий. Однако поиски предковой бактерии и реконструкция событий симбиогенеза еще далеки от завершения. Авторы новой статьи в журнале PNAS подошли к проблеме с другого конца: они смоделировали симбиогенез на примере хорошо изученной бактерии (Escherichia coli) и хорошо изученной эукариотической клетки (Saccharomyces cerevisiae). Теперь у нас есть отработанная методика получения химерных клеток, с помощью которой можно проверять, какие именно свойства предковой бактерии были необходимы для симбиогенеза.
Общий принцип, которым руководствовались авторы эксперимента, можно сформулировать так: чтобы заставить две клетки вступить в симбиоз, нужно отобрать у них что-то жизненно важное, тогда их существование по отдельности станет невозможно (рис. 2).
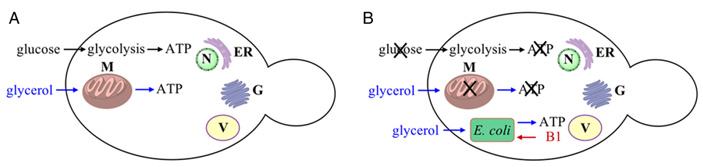
Всю работу можно условно разделить на пять шагов.
Шаг 1 — лишить кишечную палочку самодостаточности. Чтобы эндосимбиоз оказался выгодным решением для бактерии, она должна стать ауксотрофом — быть неспособной производить какое-нибудь жизненно необходимое вещество. Для многих бактерий таким веществом является тиамин (витамин B1) — кофермент в реакциях углеводного обмена. Поэтому в геноме E. coli ген биосинтеза тиамина был заменен на кассету (см. Gene cassette) с GFP (зеленым флуоресцентным белком) и геном устойчивости к антибиотику канамицину. Теперь клетки не могут выживать без внешнего источника тиамина (который они сквозь мембрану закачивают внутрь), их можно отобрать под действием антибиотика и отследить во флуоресцентный микроскоп.
Шаг 2 — сделать кишечную палочку полезной. Авторы гипотезы происхождения митохондрии из внутриклеточных паразитов полагают, что одним из ключевых белков был АТФ/АДФ-антипортер (см. Antiporter). Это белок-переносчик, который обменивает АТФ на АДФ, меняя их местами по разные стороны мембраны. У паразитической бактерии он должен работать на благо бактерии: захватывать АТФ снаружи (то есть отбирать у клетки-хозяина) и менять на отработанные АДФ бактерии. Однако этот механизм можно заставить работать и в обратную сторону, если концентрации веществ поменяются местами. При этом бактерия начнет забирать АДФ из цитоплазмы хозяина и отдавать АТФ. Так или иначе, АДФ/АТФ-антипортеры есть как у современных митохондрий, так и у внутриклеточных паразитов. У свободно живущей кишечной палочки такого белка нет, поэтому пришлось снабдить клетки E. coli плазмидой с соответствующим геном.
Шаг 3 — лишить дрожжи самодостаточности. Чтобы заставить дрожжи вступить в симбиоз, их нужно лишить энергии, то есть АТФ. Тогда единственным выходом будет получить его от кишечной палочки. Но у дрожжей, как у почти всех эукариот, есть свои митохондрии. Поэтому авторы эксперимента взяли мутантный штамм дрожжей, лишенный одного из ключевых митохондриальных генов. Такие клетки содержат митохондрии, но не получают от них энергии. Они не могут расти в среде, где из питательных веществ есть только глицерин. Однако оказалось, что и в симбиоз с E. coli они тоже не вступают.
 Читайте также:
Читайте также:

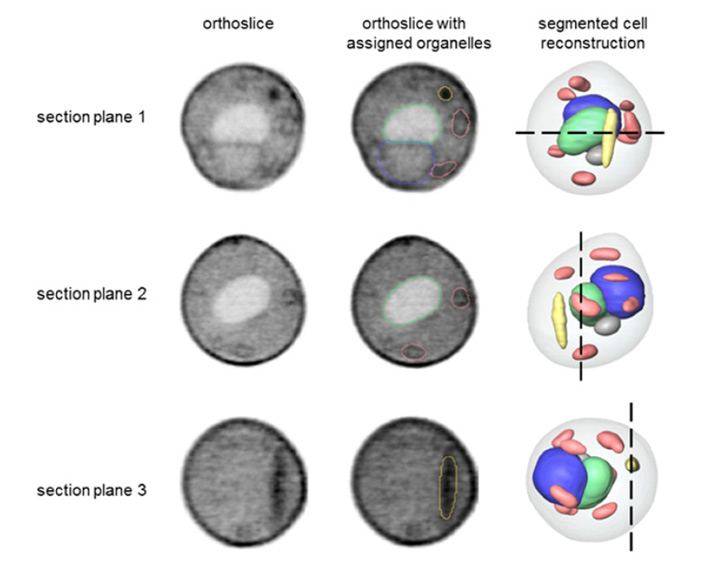
Рис. 3. Ультраструктура химерных клеток: результаты томографии под действием мягкого рентгеновского излучения. Сверху вниз — три плана в разных плоскостях. Левый столбец — просто снимок, средний столбец — снимок с выделенными органеллами, правый столбец — реконструкция клетки с обозначением плоскости среза (пунктирная линия). Изображение из обсуждаемой статьи в PNAS
Шаг 5 — убрать лишнее. В ходе эволюции митохондрия утратила большую часть ДНК (у млекопитающих, например, в ее геноме осталось лишь 37 генов). Это значит, что она становилась всё более зависимой от своей клетки-хозяина. Авторы обсуждаемой статьи попробовали воспроизвести и этот этап тоже. Для этого они удалили у клеток кишечной палочки ген биосинтеза НАД + — еще одного важного кофермента. Клетки, лишенные НАД + , так же как и их предшественники, лишенные тиамина, успешно образовывали химеры с дрожжами. И даже двойные мутанты, неспособные производить ни один из этих коферментов, также вступали в эндосимбиоз (рис. 4).
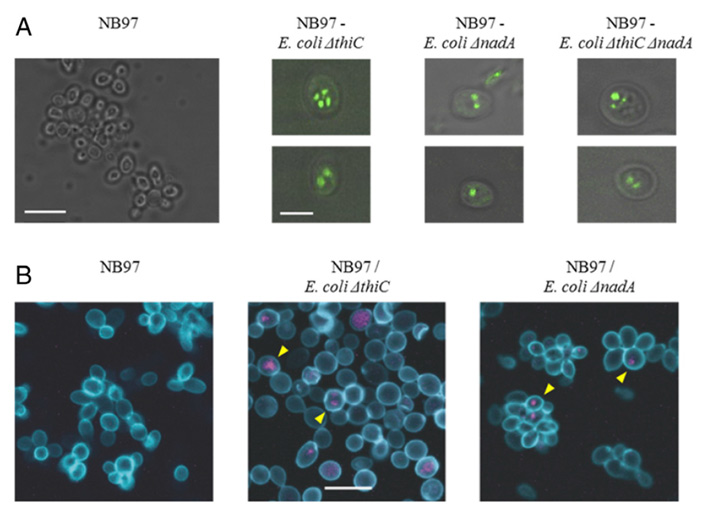
Рис. 4. Колонии химерных клеток, образованные разными штаммами кишечной палочки. А. Слева направо: контроль (клетки дрожжей), длина масштабного отрезка 10 мкм; химера с E. coli, дефицитными по тиамину, длина масштабного отрезка 5 мкм; химера с E. coli, дефицитными по НАД + ; химера с E. coli, дефицитными по обоим коферментам. Зеленым светится GFP в клетках кишечной палочки. В. Слева направо: контроль, химера с E. coli, дефицитными по тиамину и по НАД + . Дрожжи окрашены голубым (краситель FITC), бактерии — фиолетовым зондом, связывающимся с бактериальной РНК. Желтые стрелки указывают на примеры химерных клеток. Длина масштабного отрезка 10 мкм. Изображение из обсуждаемой статьи в PNAS
Перед нами — отработанная методика, с помощью которой можно моделировать ранние события эндосимбиоза. Клетки кишечной палочки, дефицитные по разным веществам, равно хорошо образуют химеры, которые воспроизводятся из поколения в поколение. Следующий шаг — поиск предельной редукции генома E. coli, возможной в данной ситуации. Авторы статьи отмечают, что удаление всего двух путей биосинтеза уже дало экономию в 7,7 тысяч пар нуклеотидов (для сравнения, весь митохондриальный геном человека составляет примерно 15 тысяч пар). Поэтому нам еще предстоит найти ту грань, на которой экономия размера генома столкнется с возможностью выживания клетки-симбионта.
Кроме того, как ехидно указывают авторы в конце текста, при таком раскладе не очень понятно, кто в этой истории настоящий паразит. Если бактерия, попавшая внутрь археи, лишь постепенно утрачивала свои метаболические пути, то возможно настоящим паразитом здесь стоит считать архею, которая потребляла энергию, производимую бактерией.
Брожение состоит из гликолиза (2 АТФ) и превращения ПВК в молочную кислоту или спирт + углекислый газ (0 АТФ). Итого 2 АТФ.
Покрыты двумя мембранами. Наружная мембрана гладкая, внутренняя имеет выросты внутрь – кристы, они увеличивают площадь внутренней мембраны, чтобы расположить на ней как можно больше ферментов клеточного дыхания.
Внутренняя среда митохондрии называется матрикс. В нем находятся кольцевая ДНК и мелкие (70S) рибосомы, за счет них митохондрии самостоятельно делают для себя часть белков, поэтому их называют полуавтономными органоидами.
- Читайте также:
2. В процессе гликолиза образовалось 84 молекулы пировиноградной кислоты. Какое число молекул глюкозы подверглось расщеплению и сколько молекул АТФ образуется при её полном окислении? Запишите два числа в порядке, указанном в задании, без разделителей (пробелов, запятых и т.п.).
3. В диссимиляцию вступило 15 молекул глюкозы. Определите количество АТФ после гликолиза, после энергетического этапа и суммарный эффект диссимиляции. Запишите три числа в порядке, указанном в задании, без разделителей (пробелов, запятых и т.п.).
4. Сколько молекул АТФ будет синтезировано в клетках эукариот на подготовительном этапе энергетического обмена, в процессе гликолиза и в процессе дыхания при окислении фрагмента молекулы крахмала, состоящего из 10 остатков глюкозы? Ответ запишите в виде последовательности трех чисел в порядке их убывания без разделителей (пробелов, запятых и т.п.).
Известно, что митохондрии – полуавтономные органоиды клеток аэробных эукариотических организмов. Выберите из приведенного ниже текста три утверждения, по смыслу относящиеся к описанным выше признакам, и запишите цифры, под которыми они указаны. (1) Митохондрии – достаточно крупные органоиды, занимают значительную часть цитоплазмы клетки. (2) Митохондрии имеют свою собственную кольцевую ДНК и мелкие рибосомы. (3) С помощью микросъемки живых клеток удалось обнаружить, что митохондрии подвижны и пластичны. (4) Клетки организмов, нуждающихся в свободном молекулярном кислороде для процессов дыхания, в митохондриях окисляют ПВК до углекислого газа и воды. (5) Митохондрии можно назвать энергетическими станциями клетки, так как выделяющаяся в них энергия запасается в молекулах АТФ. (6) Ядерный аппарат регулирует все процессы жизнедеятельности клетки, включая деятельность митохондрий.
Митохондрия – это двумембранный органоид эукариотической клетки, основная функция которого синтез АТФ – источника энергии для жизнедеятельности клетки.
Количество митохондрий в клетках не постоянно, в среднем от нескольких единиц до нескольких тысяч. Там, где процессы синтеза идут интенсивно, их больше. Также варьирует размер митохондрий и их форма (округлые, вытянутые, спиральные, чашевидные и др.). Чаще имеют округлую вытянутую форму, диаметром до 1 микрометра и длиной до 10 мкм. Могут перемещаться в клетке с током цитоплазмы или оставаться в одном положении. Перемещаются к местам, где больше всего требуется выработка энергии.
Согласно гипотезе симбиогенеза митохондрии произошли от аэробных бактерий, внедрившихся в другую прокариотическую клетку. Эти бактерии начали снабжать клетку дополнительным количеством молекул АТФ, а получать от нее питательные вещества. В процессе эволюции они потеряли автономность, передав часть своих генов в ядро и став таким образом клеточной органеллой.
В клетках новые митохондрии появляются в основном путем деления ранее существующих, т. е. они не синтезируются заново, что напоминает процесс размножения и говорит в пользу симбиогенеза.
Строение и функции митохондрии
Митохондрия состоит из
-
Читайте также:

двух мембран — внешней и внутренней,
межмембранного пространства,
внутреннего содержимого — матрикса,
крист, представляющих собой выросты в матрикс внутренней мембраны,
собственной белок-синтезирующей системы: ДНК, рибосом, РНК,
белков и их комплексов, в том числе большого количества ферментов и коферментов,
других молекул и гранул различных веществ, находящихся в матриксе.
Внешняя и внутренняя мембраны выполняют разные функции, поэтому различается их химический состав. Расстояние между мембранами составляет до 10 нм. Внешняя мембрана митохондрий по строению схожа с плазмалеммой, окружающей клетку, и выполняет в основном барьерную функцию, отграничивая содержимое органоида от цитоплазмы. Через нее проникают мелкие молекулы, транспорт крупных избирателен. В некоторых местах внешняя мембрана соединена с ЭПС, каналы которой открываются в митохондрию.
На внутренней мембране, в основном ее выростах — кристах, располагаются ферменты, образуя мультиферментативные системы. Поэтому по химическому составу здесь преобладают белки, а не липиды. Количество крист варьирует в зависимости от интенсивности процессов. Так в митохондриях мышц их очень много.
-
Читайте также:
В некоторых местах внешняя и внутренняя мембрана соединяются между собой.
У митохондрий, также как у хлоропластов, есть своя белоксинтезирующая система — ДНК, РНК и рибосомы. Генетический аппарат представляет собой кольцевую молекулу – нуклеоид, как у бактерий. Рибосомы митохондрий растений схожи с бактериальными, у животных митохондриальные рибосомы мельче не только цитоплазматических, но и бактериальных. Часть необходимых белков митохондрии синтезируют сами, другую часть получают из цитоплазмы, так как эти белки кодируются ядерными генами.
Главная функция митохондрий — снабжать клетку энергией, которая путем многочисленных ферментативных реакций извлекается из органических соединений и запасается в АТФ. Часть реакций идет с участием кислорода, в других выделяется углекислый газ. Реакции идут как в матриксе (цикл Кребса), так и на кристах (окислительное фосфорилирование).
Следует иметь в виду, что в клетках АТФ синтезируется не только в митохондриях, но и в цитоплазме в процессе гликолиза. Однако эффективность этих реакций невысока. Особенность функции митохондрий в том, что в них протекают реакции не только бескислородного окисления, но и кислородный этап энергетического обмена.
Другими словами, функция митохондрий – активное участие в клеточном дыхании, к которому относят множество реакций окисления органических веществ, переноса протонов водорода и электронов, идущих с выделением энергии, которая аккумулируется в АТФ.
Ферменты митохондрий
Ферменты транслоказы внутренней мембраны митохондрий осуществляют активный транспорт АДФ и АТФ.
В структуре крист выделяют элементарные частицы, состоящие из головки, ножки и основания. На головках, состоящих из фермента АТФазы, происходит синтез АТФ. АТФаза обеспечивает сопряжение фосфорилирования АДФ с реакциями дыхательной цепи.
Компоненты дыхательной цепи находятся в основании элементарных частиц в толще мембраны.
В матриксе находится большая часть ферментов цикла Кребса и окисления жирных кислот.
В результате активности электротранспортной дыхательной цепи ионы водорода поступают в нее из матрикса, а высвобождаются на наружной стороне внутренней мембраны. Это осуществляют определенные мембранные ферменты. Разница в концентрации ионов водорода по разные стороны мембраны приводит к возникновению градиента pH.
Энергию для поддержания градиента поставляет перенос электронов по дыхательной цепи. Иначе ионы водорода диффундировали бы обратно.
Энергия градиента pH используется для синтеза АТФ из АДФ:
АДФ + Ф = АТФ + H2O (реакция обратима)
Образующаяся вода ферментативно удаляется. Это, наряду с другими факторами, облегчает протекание реакции слева направо.
Рис. 1. Снимок химерных клеток кишечной палочки и дрожжей, сделанный с помощью конфокального флуоресцентного микроскопа. Голубым окрашены клетки дрожжей, фиолетовым — РНК кишечной палочки. Изображение из обсуждаемой статьи в PNAS
Общепринятая на данный момент теория симбиогенеза предполагает, что митохондрии в эукариотических клетках произошли от симбиотических бактерий. Однако поиски предковой бактерии и реконструкция событий симбиогенеза еще далеки от завершения. Авторы новой статьи в журнале PNAS подошли к проблеме с другого конца: они смоделировали симбиогенез на примере хорошо изученной бактерии (Escherichia coli) и хорошо изученной эукариотической клетки (Saccharomyces cerevisiae). Теперь у нас есть отработанная методика получения химерных клеток, с помощью которой можно проверять, какие именно свойства предковой бактерии были необходимы для симбиогенеза.
Общий принцип, которым руководствовались авторы эксперимента, можно сформулировать так: чтобы заставить две клетки вступить в симбиоз, нужно отобрать у них что-то жизненно важное, тогда их существование по отдельности станет невозможно (рис. 2).
Всю работу можно условно разделить на пять шагов.
Шаг 1 — лишить кишечную палочку самодостаточности. Чтобы эндосимбиоз оказался выгодным решением для бактерии, она должна стать ауксотрофом — быть неспособной производить какое-нибудь жизненно необходимое вещество. Для многих бактерий таким веществом является тиамин (витамин B1) — кофермент в реакциях углеводного обмена. Поэтому в геноме E. coli ген биосинтеза тиамина был заменен на кассету с GFP (зеленым флуоресцентным белком) и геном устойчивости к антибиотику канамицину. Теперь клетки не могут выживать без внешнего источника тиамина (который они сквозь мембрану закачивают внутрь), их можно отобрать под действием антибиотика и отследить во флуоресцентный микроскоп.
Шаг 2 — сделать кишечную палочку полезной. Авторы гипотезы происхождения митохондрии из внутриклеточных паразитов полагают, что одним из ключевых белков был АТФ/АДФ-антипортер. Это белок-переносчик, который обменивает АТФ на АДФ, меняя их местами по разные стороны мембраны. У паразитической бактерии он должен работать на благо бактерии: захватывать АТФ снаружи (то есть отбирать у клетки-хозяина) и менять на отработанные АДФ бактерии. Однако этот механизм можно заставить работать и в обратную сторону, если концентрации веществ поменяются местами. При этом бактерия начнет забирать АДФ из цитоплазмы хозяина и отдавать АТФ. Так или иначе, АДФ/АТФ-антипортеры есть как у современных митохондрий, так и у внутриклеточных паразитов. У свободно живущей кишечной палочки такого белка нет, поэтому пришлось снабдить клетки E. coli плазмидой с соответствующим геном.
Шаг 3 — лишить дрожжи самодостаточности. Чтобы заставить дрожжи вступить в симбиоз, их нужно лишить энергии, то есть АТФ. Тогда единственным выходом будет получить его от кишечной палочки. Но у дрожжей, как у почти всех эукариот, есть свои митохондрии. Поэтому авторы эксперимента взяли мутантный штамм дрожжей, лишенный одного из ключевых митохондриальных генов. Такие клетки содержат митохондрии, но не получают от них энергии. Они не могут расти в среде, где из питательных веществ есть только глицерин. Однако оказалось, что и в симбиоз с E. coli они тоже не вступают.
Рис. 3. Ультраструктура химерных клеток: результаты томографии под действием мягкого рентгеновского излучения. Сверху вниз — три плана в разных плоскостях. Левый столбец — просто снимок, средний столбец — снимок с выделенными органеллами, правый столбец — реконструкция клетки с обозначением плоскости среза (пунктирная линия). Изображение из обсуждаемой статьи в PNAS
Шаг 5 — убрать лишнее. В ходе эволюции митохондрия утратила большую часть ДНК (у млекопитающих, например, в ее геноме осталось лишь 37 генов). Это значит, что она становилась все более зависимой от своей клетки-хозяина. Авторы обсуждаемой статьи попробовали воспроизвести и этот этап тоже. Для этого они удалили у клеток кишечной палочки ген биосинтеза НАД + — еще одного важного кофермента. Клетки, лишенные НАД + , так же как и их предшественники, лишенные тиамина, успешно образовывали химеры с дрожжами. И даже двойные мутанты, неспособные производить ни один из этих коферментов, также вступали в эндосимбиоз (рис. 4)
Рис. 4. Колонии химерных клеток, образованные разными штаммами кишечной палочки. Слева направо: контроль (клетки дрожжей), химера с E. coli, дефицитными по тиамину, химера с E. coli, дефицитными по НАД + , химера с E. coli, дефицитными по обоим коферментам. А — зеленым светится GFP в клетках кишечной палочки. В — дрожжи окрашены голубым (краситель FITC), бактерии — фиолетовым зондом, связывающимся с бактериальной РНК. Желтые стрелки указывают на примеры химерных клеток. Изображение из обсуждаемой статьи в PNAS
Перед нами — отработанная методика, с помощью которой можно моделировать ранние события эндосимбиоза. Клетки кишечной палочки, дефицитные по разным веществам, равно хорошо образуют химеры, которые воспроизводятся из поколения в поколение. Следующий шаг — поиск предельной редукции генома E. coli, возможной в данной ситуации. Авторы статьи отмечают, что удаление всего двух путей биосинтеза уже дало экономию в 7,7 тысяч пар нуклеотидов (для сравнения — весь митохондриальный геном человека составляет примерно 15 тысяч пар). Поэтому нам еще предстоит найти ту грань, на которой экономия размера генома столкнется с возможностью выживания клетки-симбионта.
Кроме того, как ехидно указывают авторы в конце текста, при таком раскладе не очень понятно, кто в этой истории настоящий паразит. Если бактерия, попавшая внутрь археи, лишь постепенно утрачивала свои метаболические пути, то возможно настоящим паразитом здесь стоит считать архею, которая потребляла энергию, производимую бактерией.